
Muscle is an excitable tissue, meaning that it can be stimulated mechanically, chemically or electrically to produce an action potential. An action potential is an electrical change across a cell membrane due to changes in the conduction of ions across the membrane. Nervous tissue is also an excitable tissue. Muscle cells contain a contractile mechanism that is activated by the action potential. About 40% of the body is skeletal muscle, add another 5-10 % for cardiac and smooth muscle.
Contraction by a whole muscle
Isometric versus Isotonic
Isotonic contractions are those when the tension on a muscle remains constant but the muscle shortens as in lifting a static amount of weight. Isometric muscle contractions occur when the muscle doesn’t shorten as, for example when pushing against an immovable object like a wall. Isometric contraction differs from isotonic in that the myofibrils don’t slide over each other much as force is developed. Sliding does occur in isotonic contractions and external work is performed.
Motor unit concept
The motor nerve and all the fibers it innervates is called the motor unit. The number of fibers is dependent on the necessity for fine control. In general, small muscles that react rapidly with fine control have one nerve and only a few muscle fibers. Those muscles that do not require fine control, such as the gastrocnemius (calf muscle), may have several hundred muscle fibers per motor unit.
Summation
The contraction of individual muscle fibers is all-or-none. Therefore, any graded response must come from the number of motor units stimulated at any one time. Summation is the adding together of individual muscle twitches to make a whole muscle contraction. This can be accomplished by increasing the number of motor units contracting at one time (spatial summation) or by increasing the frequency of contraction of individual muscle contractions (temporal summation). These processes almost always occur simultaneously within normal muscle contraction. Usually, individual motor units fire asynchronously.
All motor units are not created equal. Therefore one motor unit within a particular muscle may be as much as 50 times as strong as another. Smaller motor units are much more easily excited than larger ones because they are innervated by smaller nerve fibers that have a naturally lower threshold for excitation. In spatial summation motor units are recruited by increasing the strength of the stimulus thereby increasing the strength of the contraction.
In temporal or wave summation the rapidity of each motor units contraction increases such that one contraction isn’t completely over when the stimulus for the next arrives. So the force generated in the first is added to that generated by the second, third and so on. When a muscle is stimulated at progressively greater frequencies, a frequency is finally reached at which the successive contractions fuse together and cannot be distinguished one from the other. The muscle then enters a long continual state of maximal contraction called tetany.
Muscle Fatigue
Prolonged strong contractions leads to fatigue of the muscle caused by the inability of the contractile and metabolic processes to supply adequately to maintain the work load. The nerve continues to function properly passing the action potential onto the muscle fibers but the contractions become weaker and weaker due to the lack of ATP.
Hypertrophy
Muscle hypertrophy (increase in muscle mass) is caused by forceful muscular activity. The diameters of individual fibers increase, nutrient and metabolic substances increase, mitochondria may increase, and the myofibrils also increase in size and number. Muscular hypertrophy increases the power for muscle contraction and nutritive mechanisms for motioning that increased power. Forceful muscle activity, above 75% of maximal, is necessary to produce hypertrophy which is why isometric exercise for even short periods of time can have profound effects on muscle mass. However, prolonged light exercise increases endurance, causing increases in oxidative enzymes, myoglobin, and even blood capillaries.
Atrophy
Muscle atrophy results when a muscle is not used for a length of time or is used for only weak contractions. For instance, atrophy occurs when limbs are put in casts. As little as one month of disuse can sometimes decrease the muscle size to one half normal. Damage to the nerve to a muscle results atrophy a well. If the damage is repaired in the first 3-4 months the muscle will regain full function. After four months muscle fibers will have degenerated to fibrous and fatty tissue.
Muscle Types and Mechanism of Contraction
Skeletal Muscle
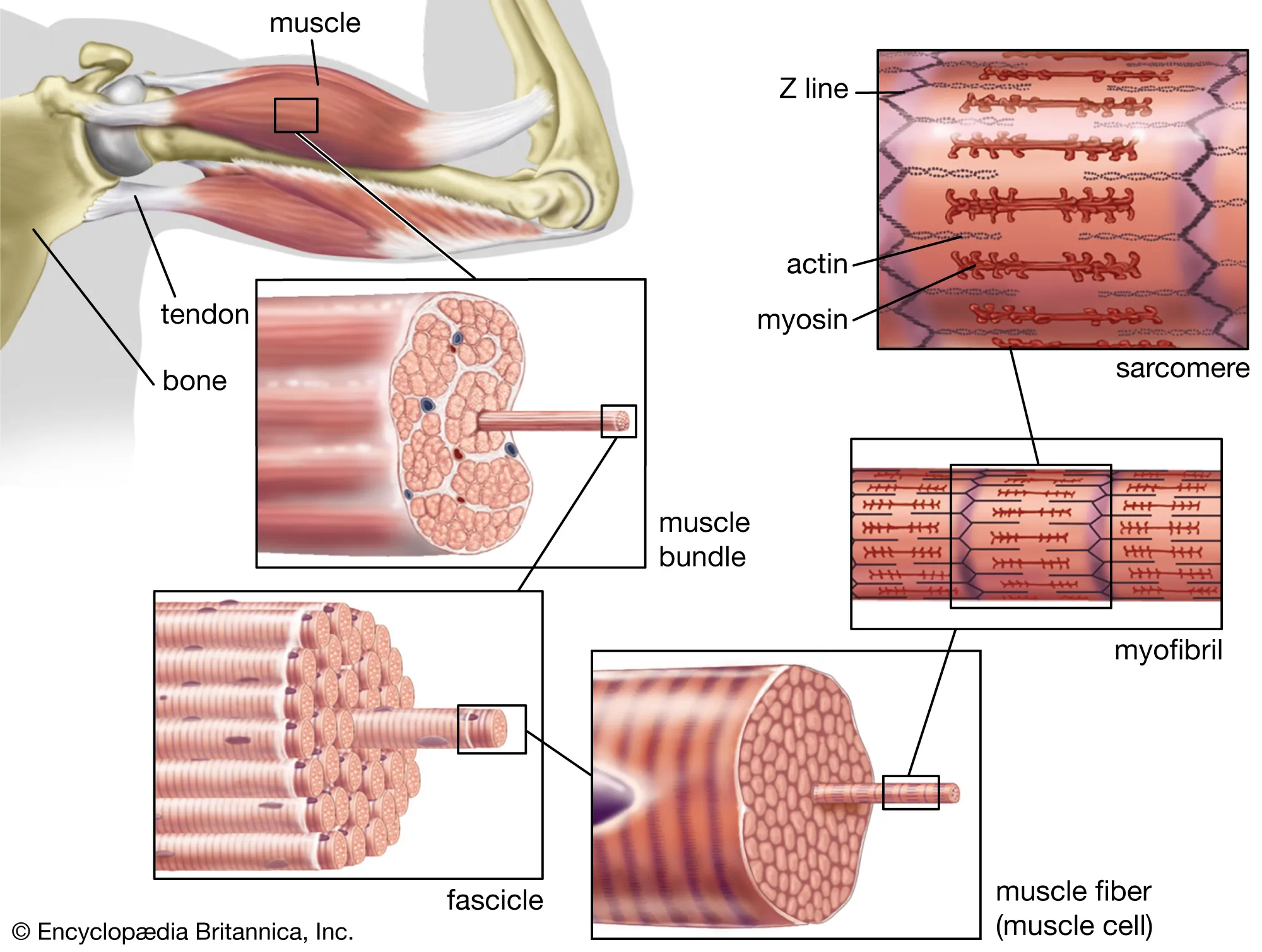
Skeletal muscle makes up most of the body’s muscle and does not contract without nervous stimulation. It is under voluntary control and lacks anatomic cellular connections between fibers. The fibers (cells) are multinucleate and appear striated due to the arrangement of actin and myosin protein filaments. Each fiber is a single cell, long, cylindric and surrounded by a cell membrane. The muscle fibers contain many myofibrils that are made of myofilaments. These myofilaments are made of the contractile proteins. The key proteins in muscle contraction are myosin, actin, tropomyosin and troponin.
Skeletal muscle fibers have differences in metabolic and contractile properties. Type I fibers are mostly found in the muscle for posture as in the long muscles of the back. These are also called red muscles because the fibers contain many mitochondia that give the muscle more of a dark reddish hue. White muscles contain mostly Type IIB fibers and are specialized for fast, fine movements as in the muscles that move the eye or some hand muscles. The differences in fiber type occur because of differences in amino acid composition of the skeletal proteins without a change in biologic activity. Various forms of the proteins can be expressed thus detemining the functional characteristics of each muscle. Changes in muscle function can be caused by alterations in activity (training), hormonal environment (steroids), or innervation. Skeletal muscle can undergo a limited regeneration in case of injury via satellite cells that are located on the periphery of the muscle fiber. These cells may be active in muscle hypertrophy as well.
Contractile Proteins
Skeletal muscle is composed of cells, called fibers, that are specialized to contract or shorten in length. Each fiber is made of smaller subunits called myofibrils that are composed of contractile proteins called myosin and actin which are responsible for muscle contraction at the molecular level. These contractile protein filaments are also called thick (myosin) and thin (actin) filaments. These filaments interdigitate such that the proteins can interact. The myosin filaments have what are called crossbridges that stick out from the filament to interact with the actin filaments during contraction. Imagine a set of golf clubs held together by their shafts with the heads radiating out around the shafts. This is a visual picture of what the thick filaments look like. Because the clubs have different length shafts the heads stick out at different places along the cluster. The myosin filaments look like this on both ends of a long filament that is made of some 200 myosin protein molecules. This structure allows the myosin filament to pull the actin filaments from both directions thus shortening the fiber.
The actin filaments are composed of two strands of protein that are woven together as one. The actin filaments are anchored to Z lines that make the boundaries of the functional unit of muscle contraction called the sarcomere. There are many sarcomeres in a muscle fiber and Z lines are continuous across muscle fibers.
Sliding Filament Theory
Muscle contraction occurs by a sliding filament mechanism whereby the sarcomeres shorten (the Z-lines come closer together) by the action of the actin filaments sliding over the myosin filaments.
Myosin filaments may look somewhat like a golf club but they are not inflexible. In fact, muscle contraction would be impossible if the myosin molecules did not have a “hinge” along the shaft that allows for a ratchet movement of the head. The force behind muscle contraction is the ratchet movement of these tiny myosin heads toward the center of their sarcomere. This ratchet movement occurs many times during a muscle contraction.
The thin filaments are actually composed of more than just actin which forms the backbone of the filament. Two other proteins are part of the thin filaments, tropomyosin and troponin. Along the actin filaments there are active sites where myosin attaches during contraction. These active sites are covered in the relaxed state by tropomyosin so that contraction cannot occur. Troponin is a complex of three submits having different affinities. One has an affinity for actin, another for tropomyosin and a third for calcium. Troponin molecules are positioned along the actin-tropomyosin filaments and act to position the tropomyosin filaments over the active sites on the actin filaments.
When calcium is present it binds to the troponin which changes in shape causing the movement of tropomyosin off the active sites so that myosin and actin can interact and muscle contraction can occur. When the active sites are uncovered the myosin heads bind to the sites which initiates a movement of the head toward the center of the sarcomere thus pulling the actin along and shortening the sarcomere. Each one of the myosin heads is thought to operate independently of the others, each attaching and pulling in a continuous alternating ratchet cycle until the calcium is removed and the active sites are covered up again.
Muscle contraction requires a great deal of energy. Energy is required to break the bond between the myosin head and the actin active sites as well as for removal of calcium from the cytoplasm by the use of a special pump within the sarcoplasmic reticulum. When the myosin head is tilted forward, after the power stroke, a binding site for ATP (the chief energy currency of the cell) is exposed. The breakdown of ATP to ADP releases the head from the actin filament and cocks it for the next ratchet power stroke.
Energy Sources
Energy is required for muscle contraction. At rest and during light exercise, muscles use lipids as their energy source. The use of carbohydrate becomes more important as the intensity of exercise increases. The breakdown of glucose to water and carbon dioxide generates energy that is transferred to regenerate phosphorylcreatine and ATP. When oxygen supplies are inadequate this process is short circuited and a metabolite (lactic acid) of one of the products builds up in the muscle. This is called anaerobic metabolism (glycolysis) and is a normal process that can occur prior to the oxidative breakdown of glucose. The lactate builds up in the muscles causing a change in pH that inhibits enzyme activity. After the exercise, an oxygen debt exists in that oxygen must be used to convert the lactate into carbon dioxide and water and replenish energy stores. Short intense exercise utilizes anaerobic metabolic mechanisms more than more sustained activities. For example, in a 100 m dash 85% of the energy is derived from anaerobic means while in a mile run only 20% is generated anaerobically.
Excitation-Contraction Coupling
Contraction in skeletal muscle begins with an action potential in the muscle fiber. This causes the release of calcium from the sacroplasmic reticulum. The action potential in the muscle fiber begins after it is excited by interaction with a large insulated (myelinated) nerve fiber. The point of contact of the nerve and muscle is called the neuromuscular junction which is normally located in the middle of the muscle fiber. Therefore an action potential initiated here spreads toward the ends of the fiber making it possible for all sarcomeres to contract at the same time. Skeletal muscle has an adaptation that allows the action potential to spread deep within the fiber. The T or transverse tubules are internal extensions of the sarcolemma that penetrate through the fiber such that action potentials in the t-tubules cause the release of calcium from the nearby sarcoplasmic reticulum in the immediate vicinity of the myofibrils.
The sarcoplasmic reticulum contains calcium ions in very high concentration that are released when the adjacent T-tubule is excited. Pumps within the walls of the sarcoplasmic reticulum return the calcium within the cytoplasm to levels below those needed to activate the contractile process.
Neuromuscular Junction
The association of the motor nerve and the muscle fiber occurs at the neuromuscular junction. Here, the neuron ends in a terminal button that contains small vesicles filled with the neurotransmitter acetylcholine. When an action potential reaches the terminal button the vesicles are released and the acetylcholine diffuses across a narrow space to bind to receptors on the muscle fiber cell membrane. When the acetylcholine binds to the receptors, the local permeability of the the muscle cell membrane is altered so that an action potential is initiated on the muscle cell. This action potential then spreads over the muscle cell membrane and T-tubule system to initiate the contractile process. An enzyme called acetylcholinesterase is present within the neuromuscular junction to break down the acetylcholine and remove the stimulus to contract.
Smooth Muscle
Smooth muscle is found in the walls of blood vessels, tubular organs such as the stomach and uterus, the iris, or associated with the hair follicles. It exists in the body as multiunit or visceral smooth muscle. It is not under voluntary control, each cell has one nucleus and it is displays automaticity in the visceral form. In multiunit smooth muscle each cell exists as a discreet independent unit that is innervated by a single nerve ending. Visceral smooth muscle exists as a sheet or bundle of fibers that are intimately connected by junctions that allow ions to flow freely and it therefore performs as a syncytium. Therefore, when one portion of visceral smooth muscle is stimulated the action potential spreads to all other fibers.

Most of the same contractile proteins are present and active in smooth muscle contraction but they are not arranged as microscopically visible parallel myofilaments as in skeletal muscle. The contractile mechanism is very similar to skeletal muscle except that the myosin of smooth muscle only interacts with actin when it has been phosphorylated. In smooth muscle calcium binds to a protein called calmodulin and the complex then interacts with an enzyme that adds a phosphate group to myosin thus activating it.
In smooth muscle, T-tubules are absent, the sarcoplasmic reticulum is poorly developed and the calcium pump is present but it is slower acting. Because of these differences in the contractile mechanism and machinery, smooth muscle takes about 30 times as long to contract and relax as does skeletal muscle and it does this while using much less energy. Elaborate neuromuscular junctions are not present in smooth muscle. Often neurotransmitter is released only in close proximity to the muscle such that the neurotransmitter, which may be acetylcholine or norepinephrine, must diffuse to the muscle cells to interact with receptors on the cell membrane. Either of these neurotransmitters may be excitatory or inhibitory depending on the receptors present on that particular smooth muscle cell. Because smooth muscle has spontaneous activity, neuronal input only serves to modify that activity rather than initiating it as in skeletal muscle. Local tissue factors, hormones and mechanical stretch can cause action potentials and thus contraction in smooth muscle. Unlike cardiac muscle, smooth muscle is capable of active regeneration after injury.
Cardiac Muscle
The heart is made of specialized muscle tissue with some similarities to both smooth and skeletal muscle. It is involuntary and mononucleate as is smooth muscle. Cardiac muscle is striated like skeletal muscle which means that it has microscopically visible myofilaments arranged in parallel with the sarcomere structure described above. These filaments slide along each other during the process of contraction in the same manner as occurs in skeletal muscle.
Cardiac muscle fibers branch and have a single nucleus per cell. Another difference in cardiac muscle is the presence of intercalated discs that are specialized connections between one cardiac muscle cell and another. These tight connections allow for almost completely free movement of ions so that action potentials can freely pass from one cell to another. This makes cardiac muscle tissue a functional syncytium. When one cell is excited the resultant action potential is spread to all of them. This is an important feature in that it allows the atrial or ventricular muscle to contract as one to forcefully pump blood. Action potentials in cardiac muscle are also specialized to maximize the pumping function of the heart. They last 10 to 30 times as long as those of skeletal muscle and cause a correspondingly increased period of contraction. Cardiac muscle has no regenerative capacity beyond early childhood.